 2016, Vol. 40
2016, Vol. 40

2. 广东省渔业生态环境重点实验室, 农业部南海渔业资源环境科学观测实验站, 农业部南海渔业资源开发利用重点实验室, 广东 广州 510300
口虾蛄(Oratosquilla oratoria)俗称虾爬子、螳螂虾及虾虎等,隶属节肢动物门(Arthropoda)、甲壳纲(Crustacen)、口足目(Stomatopoda)、虾蛄科(Squillidae),多年生大型甲壳类,广泛分布于热带及亚热带海域[1, 2, 3]。口虾蛄个体大,肉质鲜美,具有较高的经济价值[4]。其常穴居于海底泥沙砾的洞中,是凶猛的捕食性动物,主要以鱼类、虾类、贝类、蟹类和桡足类为食[4, 5, 6]。此外,作为连接海洋底层和中上层食物网的重要纽带,口虾蛄又被其他重要经济鱼类如鲈(Lateolabrax japonicus)和真鲷(Pagrosomus major)等摄食[7]。近年来,由于人类活动的影响,渔业资源衰退明显,口虾蛄的生态和经济价值也日益突显[4]。因此,研究口虾蛄食性,不仅有助于深入了解和掌握近岸海洋生态系统生物群落的结构变化和动态,而且对渔业资源的保护、可持续开发利用及生态系统的科学管理均有重要意义。
研究海洋捕食动物的营养关系是理解它们生态作用的第一步,但是想要定量收集它们的食物信息相当困难[8]。过去对水生动物食性的研究主要通过胃含物分析法进行,但此方法在食物辨认、反映食物的吸收和计算营养级等方面还存在着一些不足[9, 10]。近年来稳定同位素技术在动物学和生态学领域的应用,使得这方面研究已经取得了一些进展[11]。动物在对摄食和吸收时不造成显著的碳稳定同位素比值(δ13C)分馏,可通过δ13C值来判断动物的食物来源 [12, 13]。而氮稳定同位素比值(δ15N)在营养级传递过程中会产生富集,可用于估算消费者在食物网中的营养位置[14, 15]。稳定同位素技术的优势在于能够记录消费者所吸收的物质,而不仅仅是所吃的食物,从而反映消费者长期的摄食史[10]。此外,可利用稳定同位素混合模型,估算动物所摄食不同饵料生物的比率[11, 16]。
目前,国内外学者对口虾蛄的研究主要集中在繁殖、养殖、生物学特征和生态习性等方面[4, 17, 18, 19],对其食性定量的研究很少,均是利用胃含物分析法[5, 6],而利用稳定同位素对其食性定量的研究在国内外尚未见报道。本研究应用碳氮稳定同位素技术,对汕尾红海湾海域口虾蛄的食性进行定量分析,以期在丰富口虾蛄基础生物学资料的同时,为口虾蛄资源的保护与合理利用提供科学依据。
1 材料与方法 1.1 样品采集与处理样品来源于2015年5月在红海湾海域(114°52′~115°18′ E,22°30′~22°37′ N)进行的定点底拖网及潮间带调查。样品在冷冻的条件下被带回实验室分类鉴定和生物学测定后,进行稳定同位素样品取样。鱼类[主要由拟矛尾鳅虎鱼(Parachaeturichthys polynema)、四线天竺鲷(Apogon quadrifasciatus)、卵鳎(Solea ovata)及二长棘鲷(Parargyrops edita)等组成]取其背部白色肌肉,口虾蛄和虾类[包括墨吉对虾(Penaeus merguiensis)、细巧仿对虾(Parapenaeopsis tenella)及鲜明鼓虾(Alpheus distinguendus)等]去壳取腹部肌肉,蟹类[包括隆线强蟹(Eucrate crenata)、矛形梭子蟹(Portunus hastatoides)及远海梭子蟹(P. pelagicus)等]去壳取第一螯足肌肉,贝类[由翡翠贻贝(Perna viridis)、条纹隔贻贝(Septifer virgatus)、衣硬蓝蛤(Solidicorbula tunicata)和丽文蛤(Meretrix lusoria)组成]取闭壳肌作为分析样品。浮游动物桡足类采用大型浮游生物网由水底至水表垂直采集,用玻璃纤维滤膜(Whatman GF/F)抽滤过的海水清养2 h使其排除消化道含物,再用蒸馏水将其洗净后,在解剖镜下用解剖针将个体直接挑入到锡囊中作为分析样品。将以上稳定同位素样品在60 ℃下烘48 h至恒重,用研钵将除桡足类之外的样品研磨成粉末,装入离心管,放入干燥器中保存待测。
1.2 稳定同位素测定所采样品的稳定同位素比值在中国农业科学院农业环境稳定同位素实验室进行测定。所用仪器为Vario PYRO Cube型元素分析仪和Isoprime-100型稳定同位素比值分析仪,获得的稳定同位素比值用δ表示:
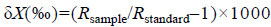
式中,X=13C或15N;R=13C/12C或15N/14N。碳和氮同位素标准样品分别采用拟箭石和大气氮。为保证实验结果的准确性和仪器的稳定性,每测试12个样品穿插1个标样进行校正。样品δ13C和δ15N值分析精度为±0.2‰。
1.3 数据处理与分析由于相同种类不同个体之间的δ13C和δ15N值较为接近,根据Phillips等[20]的方法,将它们的δ13C和δ15N值平均后进行分析。
口虾蛄及其食物的营养级(TL)通过如下公式计算:

式中,δ15Nbaseline为基线生物的δ15N(即λ=1时,δ15Nbaseline为初级生产者δ15N,而λ=2时,δ15Nbaseline为初级消费者δ15N),根据Post[21]的建议,本研究以贝类的δ15N值为δ15Nbaseline。δ15Nconsumer为口虾蛄及其食物的δ15N值,Δδ15N为营养级传递过程中δ15N的富集值,平均值约为3.4‰[21]。
本研究利用基于R程序的Bayesian稳定同位素混合模型(SIAR)计算不同饵料生物在口虾蛄食物中的贡献比率[22]。该模型是利用Bayesian方法通过不同食物和消费者的δ13C和δ15N值计算食物对口虾蛄的相对贡献。食物和口虾蛄的δ13C和δ15N值输入R语言的SIAR程序包,并经过营养富集因子校正,δ13C和δ15N值的营养富集因子分别为0.4‰±1.3‰和3.4‰±1.0‰[21],拟合得出食物对口虾蛄的贡献比率。
此外,利用Pearson相关分析检验口虾蛄个体全长和体质量与其δ13C和δ15N值的相关关系。
2 结果 2.1 口虾蛄及其食物的稳定同位素特征本研究获取口虾蛄及其食物种类共21种,其中食物种类分为5个类群(表 1)。
| 表 1 口虾蛄及其食物种类的长度和体质量 Tab. 1 Length and weight of O. oratoria and their foods |
稳定同位素测定结果表明,红海湾海域口虾蛄的δ13C值为-18.1‰~-16.3‰,均值为-17.1‰±0.5‰;δ15N值为10.9‰~13.5‰,均值为12.7‰±0.7‰(图 1)。Pearson相关分析结果表明,口虾蛄δ13C值与其个体全长和体质量间不存在显著的相关关系(r=0.060,P=0.852,n=12;r=0.545,P=0.067,n=12),其δ15N值与其个体全长间不存在显著的相关关系(r=0.383,P=0.219,n=12),但与其个体体质量间存在极显著的正相关(r=0.716,P < 0.01,n=12)(图 2)。

|
图 1 红海湾口虾蛄及其饵料生物的δ13C和δ15N值 Fig. 1 Mean±SE δ13C and δ15N values of O. oratoria and their foods in Honghai Bay |

|
图 2 口虾蛄体质量与δ13C(a)和δ15N值(b)的关系 Fig. 2 Relationships of δ13C (a) and δ15N (b) values to body weight of O. oratoria |
在口虾蛄的5类食物中,桡足类的δ13C均值最低,为-20.7‰±0.5‰;其次为贝类;鱼类及虾类的δ13C均值分别为-17.6‰±0.4‰和-16.9‰± 0.5‰;蟹类的最高。δ15N同样是桡足类的最低,贝类其次;蟹类和虾类的δ15N均值居中;而鱼类的最高,为12.6‰±0.5‰;它们的δ15N值均低于口虾蛄(图 1)。
2.2 口虾蛄的食物组成从SIAR计算结果可知,口虾蛄的主要食物为贝类,平均贡献率为38.6%;其次为蟹类和桡足类,平均贡献率分别为22.9%和16.0%;虾类的平均贡献率为13.6%;鱼类的最低,仅为8.9%(图 3)。

|
图 3 利用SIAR分析的口虾蛄食物贡献率 1.鱼类;2.虾类:3.蟹类;4.贝类;5.桡足类;条状宽度由宽变窄分别代表95%、75%和50%置信区间 Fig. 3 Results of the SIAR mixing model analysis to estimate food contribution to O. oratoria 1. fish; 2. shrimp; 3. crab; 4. bivalve; 5. copepod; widths of bars showing the 95%, 75% and 50% credibility intervals from wide to narrow |
根据δ15N值及营养级的计算公式得出,红海湾海域口虾蛄的营养级为3.01±0.22。在其5类食物当中,桡足类的营养级最低,仅为1.77±0.12;其次为贝类;蟹类和虾类的营养级分别为2.78± 0.21和2.89±0.16;鱼类的最高,为2.98±0.15;它们的营养级均低于口虾蛄。
3 讨论 3.1 口虾蛄的食物组成研究结果显示,口虾蛄的δ13C值为-18.1‰~ -16.3‰,δ15N值为10.9‰~13.5‰,δ13C和δ15N值的差值分别为1.8‰和2.6‰,与其他海洋生物相比,δ13C和δ15N值的变化范围均较大[23],表明口虾蛄的摄食选择性较大。口虾蛄的δ13C值在鱼类、虾类、贝类、蟹类和桡足类之间,而且口虾蛄的δ15N值均高于它们,因此可以判断,口虾蛄的食物组成来源于这5类生物,这与以往学者的研究结果相似[4, 5, 6]。
盛福利等[5]和徐善良等[6]通过胃含物分析法发现,虾类和鱼类是口虾蛄的重要食物组成。王春琳等[4]指出口虾蛄喜欢摄食小型虾类,其次为双壳类和小型鱼类。而本研究结果表明,贝类应为口虾蛄的主要食物,其次为蟹类和桡足类,虾类和鱼类对口虾蛄的贡献率较低。研究结果的差异可能与以下几方面原因有关:首先,从摄食行为考虑,口虾蛄寻找食物主要依赖嗅觉、化学传感器和两对触角的触觉功能,当它接近某个猎物并辨认为攻击对象后,再突然迅速地发起攻击[4]。口虾蛄在接近猎物时,猎物能否及时逃脱是其捕食成功的关键,鱼类和虾类移动能力强,桡足类和蟹类次之,而贝类的移动能力最差,因而更易被口虾蛄所摄食;其次,从食物的适口性来看,口虾蛄主要用两大颚把食物嚼碎再进食[24],鱼类有骨刺、虾类和蟹类有硬壳,口虾蛄均不易摄食[5, 24]。而贝类与其他食物相比,虽然也有硬壳,但口虾蛄当遇到带有较薄贝壳的贝类时,在用掠足基部夹碎贝壳后,连同碎片一起吞下。遇到壳很厚的贝类时,则舍去壳,设法拉出其中的肉质部食之[6]。因此,贝类可能比其他食物的适口性更好。另外,李铭五[24]研究发现,虾蛄的最适捕食指数为0.5左右,即最适于虾蛄捕食的饵料长度为虾蛄头胸甲长度或身体长度的1/3~1/2。在本研究中,鱼类的长度与虾蛄最适饵料长度相比稍长,而虾类的稍短,这可能是虾类和鱼类对口虾蛄的贡献率较低的原因。再次,口虾蛄的食物组成还受生活环境中各种饵料生物多寡的影响[6],不同海域食物数量的不同也会影响其食物贡献比率。
3.2 营养级的确定关于基线生物的选择,学者建议使用常年存在于生态系统中,而且食性简单的浮游动物或底栖动物等初级消费者。这些消费者必须与次级消费者同时长期存在,并且其稳定同位素值须能整合时间轴上的变化,而且还能反映空间上的差异[21, 25]。但是由于浮游动物的生命周期短,生长速率快[26],导致其δ15N值随时间的变化幅度大,因此作为基线生物具有其局限性。而贝类作为初级消费者,其移动距离短、寿命长,能较好地反映稳定同位素值的综合时空变化信息[21]。因此本研究选取贝类作为食物网中的基线生物来确定消费者的营养级,将其营养级定为2.0。计算结果显示,口虾蛄的营养级为3.01,表明口虾蛄为次级消费者,主要以初级消费者为食。而盛福利等[5]发现,口虾蛄的食物组成除了鱼类、虾类和蟹类外,还有一部分藻类,其食性是杂食性。但徐善良等[6]认为,口虾蛄胃含物中偶尔发现的藻类、卵子、泥砂粒等,可能是在捕食时带入口内的。
口虾蛄的食物除桡足类外,其余种类的营养级范围为2~3,介于初级消费者和次级消费者之间,表明它们除了以植物为食外,也有部分来自动物性食物。有学者报道指出,海洋蟹类的食物来源非常广泛,包括动物性食物(如线虫、多毛类、毛颚类和棘皮动物等)和植物性食物(如硅藻类、底栖藻类等),它们绝大部分为杂食性[27, 28]。本研究获取的虾类以墨吉对虾和细巧仿对虾为主,它们的食性很广,食物种类包括藻类、甲壳类和软体动物等,食物组成随发育阶段而有差别。幼虾和成虾多以肉食性为主,而仔虾则以藻类为主[29]。计算结果显示,虾类的营养级为2.89,说明它们既摄食植物,也以动物性食物为食,与本研究获取的虾类样本的发育阶段相符。在采样期间,红海湾海域桡足类的优势种主要为太平洋纺锤水蚤(Acartia pacifica),为杂食滤食性种类[30],按理它们的营养级也应为2~3,但结果仅为1.77。原因是桡足类的δ15N值受其主要食物即浮游植物的影响,而浮游植物的δ15N值受环境因子(如光照、水温和营养盐等)变化的影响[31],使得浮游植物的15N最大周转率高,稳定同位素值变化快,导致桡足类的δ15N值也相应变化得快,有时在几个月内能达6‰,跨越近2个营养级[32],从而造成桡足类的δ15N值比基线生物的低,营养级也随之低于2.0,类似的结果在其他学者的研究中也有报道[33, 34]。
本研究还发现,口虾蛄的δ15N值与其个体体质量间存在极显著的正相关,表明个体大小对口虾蛄的δ15N值有影响,较大的个体具有较高的营养层次。口虾蛄为凶猛肉食性种类,其食物组成较广杂,且小个体食谱相对较狭窄,随着其长大捕食能力增强,食谱也相应变广[6]。类似的现象在其他水生生物中也很普遍。如日本沼虾(Macrobrachium nipponensis),幼虾主要摄食轮虫和原生动物,而成虾则偏向水生昆虫和寡毛类等大型动物[35]。再如日本海鲂(Zeus faber)和欧洲无须鳕(Merluccius merluccius)等鱼类,仔鱼由于口裂较小,通常摄食微小的浮游生物,当口裂较大时,转而以其他生物为食,继而δ15N值随着食物的变化而发生改变[13, 36, 37],口裂较大的鱼类位于较高的营养层次[37, 38],δ15N值也相应较高。
| [1] | 陈丕茂, 袁华荣, 贾晓平, 等. 大亚湾杨梅坑人工鱼礁区渔业资源变动初步研究[J]. 南方水产科学, 2013, 9(5):100-108. Chen P M, Yuan H R, Jia X P, et al. Changes in fishery resources of Yangmeikeng artificial reef area in Daya Bay[J]. South China Fisheries Science, 2013, 9(5):100-108(in Chinese). |
| [2] | 徐海龙, 张桂芬, 乔秀亭, 等. 黄海北部口虾蛄体长及体质量关系研究[J]. 水产科学, 2010, 29(8):451-454. Xu H L, Zhang G F, Qiao X T, et al. Relationship between body length and body weight in mantis shrimp Oratosquilla oratoria in northern Yellow Sea[J]. Fisheries Science, 2010, 29(8):451-454(in Chinese). |
| [3] | 徐海龙, 谷德贤, 乔秀亭, 等. 渤海湾主要渔业资源长度与体质量关系分析[J]. 南方水产科学, 2014, 10(1):57-63. Xu H L, Gu D X, Qiao X T, et al. Analysis of length-weight relationship for major fishing species from Bohai Bay[J]. South China Fisheries Science, 2014, 10(1):57-63(in Chinese). |
| [4] | 王春琳, 徐善良, 梅文骧, 等. 口虾蛄的生物学基本特征[J]. 浙江水产学院学报, 1996, 15(1):60-62. Wang C L, Xu S L, Mei W X, et al. A biological basic character of Oratosquilla oratoria[J]. Journal of Zhejiang College of Fisheries, 1996, 15(1):60-62(in Chinese). |
| [5] | 盛福利, 曾晓起, 薛莹. 青岛近海口虾蛄的繁殖及摄食习性研究[J]. 中国海洋大学学报, 2009, 39(增1):326-332. Sheng F L, Zeng X Q, Xue Y. Study on propagation and feeding habits of Oratosquilla oratoria in the inshore waters of Qingdao[J]. Periodical of Ocean University of China, 2009, 39(Suppl. 1):326-332(in Chinese). |
| [6] | 徐善良, 王春琳, 梅文骧, 等. 浙江北部海区口虾蛄繁殖和摄食习性的初步研究[J]. 浙江水产学院学报, 1996, 15(1):30-36. Xu S L, Wang C L, Mei W X, et al. Preliminary studies on propagation and feeding habits of Oratosquilla oratoria in northern Zhejiang sea areas[J]. Journal of Zhejiang College of Fisheries, 1996, 15(1):30-36(in Chinese). |
| [7] | 杨纪明. 渤海鱼类的食性和营养级研究[J]. 现代渔业信息, 2001, 16(10):10-19. Yang J M. A study on food and trophic levels of Baohai Sea fish[J]. Modern Fisheries Information, 2001, 16(10):10-19(in Chinese). |
| [8] | Kiszka J J, Méndez-Fernandez P, Heithaus M R, et al. The foraging ecology of coastal bottlenose dolphins based on stable isotope mixing models and behavioural sampling[J]. Marine Biology, 2014, 161(4):953-961. |
| [9] | Bitterlich G, Gnaiger E. Phytoplanktivorous or omnivorous fish? Digestibility of zooplankton by silver carp, Hypophthalmichthys molitrix(Val.)[J]. Aquaculture, 1984, 40(3):261-263. |
| [10] | Lajtha K, Michener R H. Stable Isotopes in Ecology and Environmental Science[M]. Oxford:Blackwell, 1994:138-157. |
| [11] | Fry B. Stable Isotope Ecology[M]. New York:Springer, 2006. |
| [12] | DeNiro M J, Epstein S. Influence of the diet on the distribution of carbon isotopes in animals[J]. Geochimica et Cosmochimica Acta, 1978, 42(5):495-506. |
| [13] | Fry B, Sherr E B. δ13C measurements as indicators of carbon fiow in marine and freshwater ecosystems[J]. Contributions in Marine Science, 1984, 27(8):13-47. |
| [14] | Cabana G, Rasmussen J B. Comparison of aquatic food chains using nitrogen isotopes[J]. Proceedings of the National Academy of Sciences of the United States of America, 1996, 93(20):10844-10847. |
| [15] | Vander Zanden M J, Rasmussen J B. Primary consumer δ13C and δ15N and the trophic position of aquatic consumers[J]. Ecology, 1999, 80(4):1395-1404. |
| [16] | Doi H. Spatial patterns of autochthonous and allochthonous resources in aquatic food webs[J]. Population Ecology, 2009, 51(1):57-64. |
| [17] | 王波, 张锡烈, 孙丕喜. 口虾蛄的生物学特征及其人工苗种生产技术[J]. 黄渤海海洋学报, 1998, 16(2):64-73. Wang B, Zhang X L, Sun P X. On biological characters and artificial seedling-rearing techniques of mantis shrimp(Oratosquilla oratoria)[J]. Journal of Oceanography of Huanghai & Bohai Seas, 1998, 16(2):64-73(in Chinese). |
| [18] | Kodama K, Horiguchi T, Kume G, et al. Effects of hypoxia on early life history of the stomatopod Oratosquilla oratoria in a coastal sea[J]. Marine Ecology Progress Series, 2006, 324(8):197-206. |
| [19] | Kodama K, Shimizu T, Yamakawa T, et al. Reproductive biology of the female Japanese mantis shrimp Oratosquilla oratoria(Stomatopoda) in relation to changes in the seasonal pattern of larval occurrence in Tokyo Bay, Japan[J]. Fisheries Science, 2004, 70(5):734-745. |
| [20] | Phillips D L, Newsome S D, Gregg J W. Combining sources in stable isotope mixing models:alternative methods[J]. Oecologia, 2005, 144(4):520-527. |
| [21] | Post D M. Using stable isotopes to estimate trophic position:Models, methods, and assumptions[J]. Ecology, 2002, 83(3):703-718. |
| [22] | Parnell A C, Inger R, Bearhop S, et al. Source partitioning using stable isotopes:coping with too much variation[J]. PLoS One, 2010, 5(3):e9672. |
| [23] | 蒋日进, 章守宇, 王凯, 等. 枸杞岛近岸海域食物网的稳定同位素分析[J]. 生态学杂志, 2014, 33(4):930-938. Jiang R J, Zhang S Y, Wang K, et al. Stable isotope analysis of the offshore food web of Gouqi Island[J]. Chinese Journal of Ecology, 2014, 33(4):930-938(in Chinese). |
| [24] | 李铭五. 虾蛄食性的研究[J]. 海洋渔业, 1988(6):288. Li M W. Study on feeding habits of mantis shrimp[J]. Marine Fisheries, 1988(6):288(in Chinese). |
| [25] | Vander Zanden M J, Rasmussen J B. Variation in the δ15N and δ13C trophic fractionation:implications for aquatic food web studies[J]. Limnology and Oceanography, 2001, 46(8):2061-2066. |
| [26] | 郑重, 李少菁, 连光山. 海洋桡足类生物学[M]. 厦门:厦门大学出版社, 1992. Zheng C, Li S J, Lian G S. Copepod Biology of Marine[M]. Xiamen:Xiamen University Press, 1992(in Chinese). |
| [27] | 戴爱云, 杨思谅, 宋玉枝, 等. 中国海洋蟹类[M]. 北京:海洋出版社, 1986. Dai A Y, Yang S L, Song Y Z, et al. Crabs of theChina-Seas[M]. Beijing:China Ocean Press, 1986(in Chinese). |
| [28] | 黄美珍. 福建海区拥剑梭子蟹、红星梭子蟹和锈斑蟳的食性与营养级研究[J]. 台湾海峡, 2004, 23(2):159-166. Huang M Z. Study on feeding habit and nutrient level of Portunus argentatus, P. sanguinolentus and Charybdis feriatu in Fujian sea area[J]. Journal of Oceanography in Taiwan Strait, 2004, 23(2):159-166(in Chinese). |
| [29] | 李太武. 海洋生物学[M]. 北京:海洋出版社, 2013. Li T W. Marine Biology[M]. Beijing:China Ocean Press, 2013(in Chinese). |
| [30] | 李开枝, 谭烨辉, 黄良民, 等. 珠江口浮游桡足类摄食研究[J]. 热带海洋学报, 2012, 31(6):90-96. Li K Z, Tan Y H, Huang L M, et al. Feeding of planktonic copepods in the Pearl River Estuary[J]. Journal of Tropical Oceanography, 2012, 31(6):90-96(in Chinese). |
| [31] | Gu B. Variations and controls of nitrogen stable isotopes in particulate organic matter of lakes[J]. Oecologia, 2009, 160(3):421-431. |
| [32] | Syväranta J, Hämäläinen H, Jones R I. Within-lake variability in carbon and nitrogen stable isotope signatures[J]. Freshwater Biology, 2006, 51(6):1090-1102. |
| [33] | Kürten B, Painting S J, Struck U, et al. Tracking seasonal changes in North Sea zooplankton trophic dynamics using stable isotopes[J]. Biogeochemistry, 2013, 113(1-3):167-187. |
| [34] | 宁加佳, 杜飞雁, 王雪辉, 等. 基于稳定同位素的六指马鲅(Polynemus sextarius)食性特征分析[J]. 海洋与湖沼, 2015, 46(4):758-763. Ning J J, Du F Y, Wang X H, et al. Feeding habits of sixfinger threadfin Polynemus sextarius revealed in stable isotope analysis[J]. Oceanologia et Limnologia Sinica, 2015, 46(4):758-763(in Chinese). |
| [35] | 刘军, 龚世园, 何绪刚, 等. 武湖日本沼虾食性的研究[J]. 淡水渔业, 2005, 35(1):25-28. Liu J, Gong S Y, He X G, et al. Study on feeding habits of Macrobrachium nipponensis[J]. Freshwater Fisheries, 2005, 35(1):25-28(in Chinese). |
| [36] | Owens N J P. Natural variations in 15N in the marine environment[J]. Advances in Marine Biology, 1987, 24:389-451. |
| [37] | Karpouzi V S, Stergiou K I. The relationships between mouth size and shape and body length for 18 species ofmarine fishes and their trophic implications[J]. Journal of Fish Biology, 2003, 62(6):1353-1365. |
| [38] | Keast A, Webb D. Mouth and body form relative to feeding ecology in the fish fauna of a small lake, Lake Opinicon, Ontario[J]. Journal of the Fisheries Research Board of Canada, 1966, 23(12):1845-1874. |

 , WANG Xuehui1, 2, GU Yangguang1, 2, WANG Lianggen1, 2, LI Yafang1, 2
, WANG Xuehui1, 2, GU Yangguang1, 2, WANG Lianggen1, 2, LI Yafang1, 2 2. Guangdong Provincial Key Laboratory of Fishery Ecology and Environment, Scientific Observing and Experimental Station of South China Sea Fishery Resources and Environments, Key Laboratory of South China Sea Fishery Resources Development and Utilization, Ministry of Agriculture, Guangzhou 510300, China